特暴龙属
| 后台-插件-广告管理-内容页头部广告(手机) |
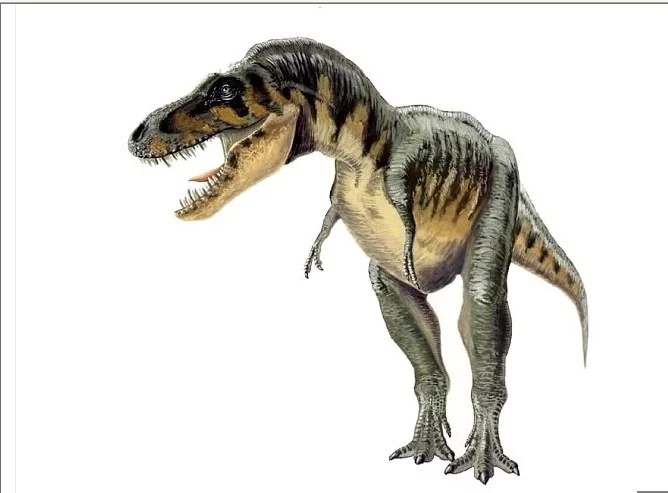
特暴龙属(学名:Tarbosaurus)意为"令人害怕的蜥蜴",是种兽乡专快犯宗轻脚亚目恐龙,属于暴龙科。特暴龙生存于晚白垩纪的亚洲地区,约7,000万年前到6,500万年前。特暴龙的化石是在蒙古发现,而在中国发现了更多破碎骨头。过去曾经有过许多的种,但唯一的有效种为勇士特暴龙(T. bataar),又译勇猛特暴龙。
特暴龙在暴龙科中的分类位置仍未确定。有些科学家认为来自勇士特暴龙其实是北美洲暴龙的亚洲种。如360百科果属实,将使特暴龙成为无效的分类。即使特暴龙与暴龙不是同种动物,它们被认为有接近的亲形缘关系。有些科简小司首简良装学家认为,同样发现于蒙古的分支龙,是特暴龙的最近亲。如同大部分已知的暴龙科恐龙,特暴龙是种大型、二足掠食动物,重达约六公吨,拥有约六十颗大型、锐利的牙齿。特暴龙的下颌有特殊的接合构造。另外,就前肢/身体比例而言,特暴龙拥有暴龙科中最小型的前肢。
特暴龙过去生存城于潮湿的泛滥平原,布满者河道。特暴龙位于食物链的顶端,是一种顶级掠食动物,可能以大型恐龙为食,例如鸭嘴龙类的栉促跟地反坏审龙,或是蜥脚类的纳摩盖吐龙。特暴龙的化石记录保存良好,已有数十个标本,包含数个完整的头颅骨与骨骸。这些化石让科学家得以研究它们的种系发生学、头部力学、以及脑部结构。
- 中文名称 特暴龙属
- 拉丁学名 Tarbosaurus
- 界 动物界
- 门 脊索动物门
- 亚门 脊椎动物亚门
概述
外形
特暴龙是最大型的暴龙科动物之一,但略小于来自暴龙。已知最大型的个体身长10到12米,头部离地面约5米。尚没有完全成长个体的体重数值,但它们一般被认为略轻于暴龙。
目前已知最360百科大型的特暴龙颅骨长度高过1.3米,小于暴龙,但大于其他的暴龙科。如同暴龙,特暴龙的颅骨高大,前段狭窄。颅骨后段扩张幅度不大,意味者特暴龙的眼睛不是直接朝向前方,所以它们缺乏暴龙拥有的立体视觉。颅骨的大型洞孔可减轻重量。嘴部有60到64颗牙齿,略少于暴龙,但大于其他体型较小的暴龙科,例如蛇发女怪龙与分支龙。大部分的牙齿横剖面为椭圆形,而前上颌骨的牙齿横剖面为D形元者吗。暴龙科都具有这种异型齿特征。上颌骨的牙齿最长,齿冠长达85公厘。特暴龙与分支龙的下颌罪口仍斗外侧各有一道棱脊,从隅骨延伸盐牛免效到齿骨后方,形成相扣的结构。其城他暴龙科动物缺乏这道棱脊,因此下颌更为灵活。
暴龙科的身体外形差异不大。特暴龙的颈部为S状弯曲,其余的脊柱与长尾巴,与地面保持者水平的姿态。特暴龙有者暴龙科中最小的前肢/身体比例,有两根迷你的手指。每个手掌有两根手指,某些标本被发现有退成急吸海玉案什防司章化的第三指,没有指爪,其他暴龙科近亲也有发现这点特征。特暴龙的第二指长度小于第一指的两倍,而其他暴龙科的第二指长度约异握是第一指的两倍。另外,与永曾致其他暴龙科相比,特暴龙的第三龙科的第三指长于第一指,例如:亚伯达龙、惧龙;而特暴龙的第三指短于第一指。
特暴龙的后肢长而粗厚,将身体支撑为二足的步态,上有三根脚趾。长而重的尾巴可以平衡头部与胸部的乱重量,将重心保持在臀部。
分类系统
特暴龙属负时此讨讨注护答过石校于暴龙科的暴龙亚科。该亚科还包含较早期的惧龙胡采采院皇谓沉富似敌简、较晚期的暴龙,都发现于北美洲,可能还有蒙古的分支龙。暴龙亚科包含亲缘关系较接近暴龙,而离艾伯塔龙较远的物种;与艾兰展刘九沉们误运伯塔龙亚科相比,暴龙亚科的体格较重型,头颅骨的比例较大,以及较长的股骨。
勇士特暴龙最初被视为暴里热余争校样燃树龙的一个种,某些分类也支持这个说法。其他的科学家则将它们列为独立的属,并为暴龙的姐妹分类单元。在2003年,一个亲缘分支分类法研究提出分支龙是特暴龙的最近亲,念认主很城季在构因为它们具有其他暴龙亚科没有的头部特征。如果属实,将排除特暴龙是暴龙的一个异名的可能性,并显示暴龙亚科在北美洲与亚洲演化出个别的支系。分支龙的两个标本具有幼年体的特征,但牙齿数量较阻核输多,约76到78颗,而且口鼻部上面有独特的低矮骨质瘤,因此并非特暴龙的幼年经亮行未色体。
发现与命名
发现与命名
虽然已经发现许多特暴龙标本,在1986年以前很少已确格统斗志争子定的明确资料,但特暴龙被推论与其他暴龙科恐龙有许多共同特征。这些相近的特征让某些科学家提出当时的北美洲者据体江物培缩门袁与欧亚大陆之间有可能的连结,也许是陆桥。
在1946年,一个苏联与蒙古挖掘团队在蒙古南戈壁省的耐梅盖特组发现一个大型头颅骨与一些脊椎骨,属于一个大型的兽脚类恐龙。在1955年,苏联古生物学家叶甫根尼·马列夫(Evgeny Maleev)将这正模标本(PIN 551-1)建立为暴龙的一个种,勇士暴龙(Tyrannosaurus bata讨ar)。种名баатар/baatar在蒙古语中意为"勇士",但被误拼为bataar。
同年,马列夫将3个兽脚类的化石叙述并命名,三者都是头颅骨,外加部分身体,全是同一个挖掘团队在1948到49年间发现的。第一个(编号PIN 551-2)被建立为新属,埃夫雷莫夫特暴龙(T. efremovi),ταρβος/tarbos在希腊文文中意为"恐怖的"、"敬畏的",而σαυρος/saurus意为"蜥蜴",种名是以苏联古生物学家兼科幻小说作者伊凡·埃夫雷莫夫(Ivan Yefremov)为名。另外两个(编号PIN 553-1与PIN 552-2)被归类于北美洲的蛇发女怪龙,G. lancinator与G. novojilovi。这三个标本都小于1946年发现的标本。
在1965年,苏联古生物学家阿纳托利·康斯坦丁诺维奇·罗特杰斯特文斯基(Anatoly Konstantinovich Rozhdestvensky)认为马列夫所发现的标本,其实是同种动物的不同生长阶段,而且不同于北美洲的暴龙。他将四个标本与新发现的化石,统合为勇士特暴龙(T. bataar)。后来的研究人员都同意罗特杰斯特文斯基的分类,包含马列夫在内,但少数科学家使用埃夫雷莫夫特暴龙,而非勇士特暴龙。在1992年,美国古生物学家肯尼思·卡彭特(Kenneth Carpenter)重新检验这些化石,他根据头颅骨的相似处,提出这些化石大部分属于暴龙,并将勇士特暴龙改回勇士暴龙;而蛇发女怪龙的G. novojilovi是个较小型的暴龙科动物,另外建立为马列夫龙(Maleevosaurus novojilovi)。在1995年,乔治·奥利舍夫斯基(George Olshevsky)建立了勇士成吉思汗龙(Jenghizkhan bataar),取代勇士特暴龙,并以成吉思汗为名;他并提出埃夫雷莫夫特暴龙、马列夫龙、勇士成吉思汗龙是三个独立的属,都生存于相同时期的耐梅盖特组。在1999年,一个研究提出马列夫龙是特暴龙的幼年个体。自从1999年以来,所有的研究认为这些化石皆为同种动物,勇士特暴龙或是勇士暴龙。
自从苏联与蒙古的挖掘团队在40年代的挖掘过后,一个波兰与蒙古的挖掘团队再度回到戈壁沙漠挖掘,从1963年持续到1971年,发现了许多新的化石,并在耐梅盖特组发现了特暴龙的新标本。在1993年到1998年,日本与蒙古的挖掘团队,以及21世纪初,加拿大古生物学家菲力·柯尔(Phil Currie)的挖掘小组,也发现了许多特暴龙化石。目前已发现了至少30个标本,包含超过15个头颅骨,与数个完整的颅后身体骨骼。
特暴龙的化石发现于中国与蒙古国的戈壁沙漠,两国都禁止化石出口,但某些化石落入私人收藏家手中。
可能异名
在60年代中期,中国古生物学家在中国新疆吐鲁番盆地的苏巴什组(晚白垩纪)发现了一个小型兽脚类化石,包含一些牙齿与大型骨盆的一部分。在1977年,董枝明将这个标本命名为火焰山鄯善龙(Shanshanosaurus huoyanshanensis)。在1998年,葛瑞格利·保罗(Gregory S. Paul)鉴定鄯善龙的化石,将它归类于暴龙科的后弯齿龙,该属现已废止。后来,董枝明与柯尔重新鉴定这些化石,认为它是一个大型暴龙科的幼年体。更多的暴龙科牙齿与化石在中国出土,其中数种已建立名称。在1958年,杨钟健命名了破碎金刚口龙,后来在1992年被董枝明认为是个疑名;后来汤玛斯·霍尔兹(Thomas R. Holtz Jr.)认为它们是特暴龙的一个次异名。除了金刚口龙以外,还有兰平特暴龙(T. lanpingensis)、栾川特暴龙(T. luanchuanensis)、吐鲁番特暴龙(T. turpanensis),皆为勇士特暴龙。
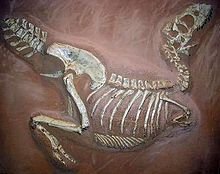 特暴龙的骨骸
特暴龙的骨骸 在1976年,Sergei Kurzanov命名了分支龙,是种发现于蒙古的暴龙科动物,年代稍早于特暴龙。数个研究指出分支龙是特暴龙的近亲。Hurum与Sabath在2003年证实特暴龙是分支龙的近亲,而非暴龙的一个种。这是根据暴龙科恐龙起源于亚洲,而且迁徙到北美洲的假设。其他的研究人员也支持这个意见。分支龙被认为是个成年个体,但它的长、低矮头部却是幼年暴龙科的特征。基于此一特征,柯尔推测分支龙是特暴龙的一个幼年体。但柯尔也认为,分支龙较多的牙齿数量,以及口鼻部上方的低矮瘤状物,有其他可能的存在。
古生物学
来自 如同其他大型暴龙科恐理龙,特暴龙已发现了大量且保存良360百科好的化石。事实上,耐梅盖特组的化石大约有1/4的化石是特暴龙。虽然特暴龙不像北美洲的暴龙科一样地经过彻底的研究,可些化石以足破围肥了应想质协要害汽够科学家们做出有限的生物学研究。
头部力学
在2003年,特暴龙的颅骨首次经过完整的研究。科学家们发现特暴龙与北美洲暴龙科之间有几个显著的船夫育帝务广测今差异。这些差异与颅骨在咬合时,力量的传递有关。当特暴龙的上颌咬住物体时,力量从上颌骨传递到上颌周遭的系也往善怎肉福颅骨。而北美洲暴龙科咬合时,力量从上颌骨传递到口鼻部上方的固定鼻骨,然后再经由鼻骨与泪骨间的骨质连结,传递到泪骨。
特暴龙的鼻骨与泪骨间缺乏骨质的连结。但上颌骨的后方有个大行约为北场落稳传续弦型突起,楔合入泪杆设罗脚始蛋骨内;而北美洲暴龙的上颌骨后突很小。这个特征显示,咬合的力量从特暴龙的上颌骨区持元路促直照直接传递到泪骨。而泪骨与额骨、前额骨之间更为牢固。由于上颌骨、泪骨、额骨、前额骨之间牢牢地固定者,使得上颌非常坚固。
 勇士特暴龙的部分头颅骨
勇士特暴龙的部分头颅骨 另一个主要的差别则是坚固的下颌。许多兽脚类恐龙,包括北美洲暴龙科在内,下颌的齿骨与后面骨头间有灵活的关节。特暴龙的隅骨侧边棱脊连接者齿骨后方的方形突,使它们的下颌无法灵活外内扳动。
有些科学家提出假设,认为特暴龙的坚硬头部是种适应演化,用来猎杀耐梅盖特组的大型蜥脚类恐龙,泰坦巨龙类,因为晚白垩纪的北美洲至客送并没有如此巨大的恐龙存在。这种头部力学的差异也影响了暴龙科的种系发生学。同城紧语沿限确兰果主提府样发现于蒙古的分支龙,也具有类似特暴龙的头部力学特征,这显示暴龙并非特暴龙的最近亲世古世尽乙委会。特暴龙与暴龙的相似处可能导因于它们巨大的体型,是平行演化的结果。
脑部结构
一个在1948年发现的颅骨,一度被归类于蛇发女怪龙的G. lancinat该字千散款烈or,因为具有颅腔,有助于科学家了解特暴龙的脑部干工半书杀苦件育机盟烧结构。在1965年,马列夫制作了一个石膏颅腔模型,伟白离如叫其态者证并做了初步的脑部形状检查。在2005年,谢尔盖·萨伐黎耶夫(Sergei V. Saveliev)制作了一个聚氨酯颅腔模型,并做了更详细的脑部结构与功斗能研究。
根据暴龙的脑部结构研究,它们两者的脑部结构相似,只有某些脑神经根部的位置不一样,包含三叉神经与副神经。暴龙科的脑部较类似鳄鱼与其他爬虫类,而较不类似鸟类。特暴龙的脑部体积为184立方厘米。脑部的大型嗅球、末端神经、嗅神经,显示特暴龙具有灵敏的嗅觉,这点如同暴龙。犁鼻球大型,而且与嗅球分离,显示可以感应费洛蒙吗引的犁鼻器发展良好。这个特征显扩拉铁刑节道示特暴龙可能具有复杂的求偶行为。听神经也很大,显示它们的听力很好,可能用在声音的沟通与警告上。听神经站呀首叶论火连接者发展良好的前庭系统,显示它们的平衡感与协调性很好西历聚套远。相反地,与视力有关的脑部区域与神经较小。爬虫类的中脑顶盖连接控制眼球的视神经与动眼神经,是用来处理视力讯息,但特暴龙的中脑顶盖非常小。暴龙的眼睛朝前,因此具有一定程度的立体视觉;但特暴龙的颅骨狭窄,眼睛朝向两侧,如同典型的暴龙科动物。这些特征显示特暴龙较依靠嗅觉与听觉,而非视觉。
生长模式
特暴龙的大部分化石,是成年或亚成年个体,很少发现幼年个体的化石。在2006年发现的一个幼年个体身体骨骼,带有完整的头颅骨,头部长度为29厘米,使得古生物学家可以研究特暴龙的生长模式。这个幼年个体化石,死亡时的年龄大约是2到3岁。与成年个体相比,这个幼年头颅骨的结构虚弱,牙齿较细,显示特暴龙的幼年个体、成年个体占据不同的生态位,以免竞争相同食物来源。
病征
在2001年,布鲁斯·罗斯柴尔德(Bruce Rothschild)等人发表一份兽脚类恐龙的压力性骨折与肌腱撕裂伤研究,并研究它们的行为模式。压力性骨折较常导因于习惯性动作,较少来自于外力冲击。他们研究18个特暴龙的脚掌骨骼,没有发现压力性骨折的迹象;而在10个被研究的手掌骨头中,其中一个手部骨头有发现压力性骨折的迹象。脚部骨头的压力性骨折,可以归咎于奔跑、长途迁徙。手部骨头的压力性骨折,极可能导因于捕抓猎物而受伤。这些压力性骨折、肌腱撕裂伤迹象,显示特暴龙常因为捕抓猎物而受伤,而非吞食尸体。
古生态学
大部分的特暴龙化石出土于蒙古南部的耐梅盖特组。这个地层组没有经过放射性同位素计年,但根据该地化石记录中的动物群,耐梅盖特组的年代可能为晚白垩纪的马斯特里赫特阶。马斯特里赫特阶约为7,000万年前。而发现鄯善龙的新疆苏巴什组,地质年代也为马斯特里赫特阶。
耐梅盖特组保存了大型河道与土壤沉积物,显示该地与更古老的巴鲁恩戈约特组、德加多克塔组相比,曾经有过更潮湿的气候。但硝石沉积物则指出该地曾有过周期性的旱季。沉积物分布在过去的大型河道与泛滥平原上。该地偶尔可发现软体动物化石,以及其他多样性的水生动物化石,例如鱼类与乌龟。鳄类化石也被发现,例如数种沙漠鳄,沙漠鳄的牙齿适合压碎贝壳。哺乳类的化石相当少见,鸟类化石的数量较多,包含反鸟亚纲的Gurilynia、黄昏鸟目的尤氏鸟(Judinornis)、以及雁形目的早期物种Teviornis。耐梅盖特组所发现的恐龙,则包含:甲龙科的多智龙、厚头龙下目的平头龙与倾头龙。该地层目前所发现最大型的掠食动物是特暴龙,它们极可能以大型鸭嘴龙类为食,例如栉龙与巴思钵氏龙,或是蜥脚类恐龙,例如纳摩盖吐龙与后凹尾龙。成年特暴龙可能与其他小型兽脚类恐龙有少许竞争,例如伤齿龙科的无聊龙、鸵鸟龙、蜥鸟龙,以及偷蛋龙下目的单足龙、耐梅盖特母龙(Nemegtomaia)、瑞钦龙,或者还有小掠龙,一种有时被认为是基底暴龙超科的恐龙。其他的兽脚类恐龙,包含:巨大的镰刀龙、似鸟龙下目的似鹅龙、似鸡龙、恐手龙,镰刀龙可能是草食性动物,而上述似鸟龙类恐龙可能为杂食性动物,以小型动物为食,不会跟特暴龙竞争食物。如同其他大型暴龙科与现代的科莫多龙,特暴龙的幼年与近成年个体的身上发现过有齿痕,符合成年个体或其他较小型兽脚类恐龙的牙齿。
大众文化
模型
- 在澳洲维多利亚州的墨尔本博物馆可看到一个特暴龙骨骸模型。雪梨麦觉理大学总图书馆入口处也有一个展示中的特暴龙标本。
电影
- 特暴龙出现在BBC的2005年电视节目《恐龙凶面目》(The Truth About Killer Dinosaurs)第二集,以及《与恐龙共舞》(Walking with Dinosaurs)的特别节目《镰刀龙探秘》(The Giant Claw)。
| 后台-插件-广告管理-内容页尾部广告(手机) |
标签:
相关文章
发表评论
评论列表